"起床!量體溫了!"麻煩給七千萬年前恐龍發條信息-來源:科學大院
 志願者在為居民測體溫(圖片來源:新華網肖和勇攝)
志願者在為居民測體溫(圖片來源:新華網肖和勇攝)
"起床!量體溫了!"麻煩給七千萬年前恐龍發條信息
-新浪科技綜合-來源:科學大院-2020年03月04日09:37
由於最近的新冠疫情,量體溫成為了人們的日常。人類的正常體溫在36到37攝氏度之間,那麼你有沒有想過,已經滅絕的恐龍的體溫是多少?
“既然想知道恐龍的體溫,那我們就直接測一測就好了!”近期,來自耶魯大學和劍橋大學的科學家們表示,我們隔著七千萬年,也能把恐龍的體溫給測出來!
科學家們用了什麼法寶呢?
恐龍到底是啥動物?
體溫測量之所以可行,是因為人類是典型內溫性動物。內溫性動物可以自主地把自身體內環境溫度穩定在一個相對固定範圍,受環境影響不大。每種內溫性動物的恆定體溫都不大相同,整體而言新陳代謝更快的動物體溫更高,比如一些小型鳥類,體溫可達40攝氏度以上。
等等,我聽過恆溫動物,冷血動物,什麼是內溫性動物?有內溫性動物,那有外溫性動物嗎?
內溫性是指一種生物可以保持其身體新陳代謝有利的溫度,並且主要依靠自身產生的熱量維持體溫,而不是主要依靠環境的熱量來維持,典型的代表是大部分哺乳動物和鳥類。
外溫性是指一種生物主要依靠外部熱源的熱量來調節體溫,它們的體溫因此也多隨著環境溫度變化,蜥蜴、鱷魚和大部分龜鱉類都是外溫性生物。
除了內溫性和外溫性動物,還有中溫性動物和巨溫性動物。
中溫性生物的體溫調節能力介於兩者之間,往往可以通過自身產的熱量維持體溫高於環境溫度,或者維持在某個區間。但不能像內溫性生物那樣將體溫維持到一個很高,並且非常穩定的程度。現生動物的代表很少,包括棱皮龜、單孔類哺乳動物等。
而巨溫性也稱慣性恆溫性。巨溫性的動物由於體型很大,導致體表表面積相對變小,散熱能力相對小體型動物下降。散熱能力下降導致它們即使新陳代謝能力不如典型的內溫性動物,但體溫也可以維持在一個很高的水平。
好了,我們回到恐龍這個話題。
研究現生動物是否是內溫性動物,只需要溫度計就可以解決。而像霸王龍這樣的已經從地球上滅絕的動物是否為內溫性動物,就很難直接測量了。分類學研究認為,恐龍與現生爬行動物(尤其是鱷類)是近親,同時又是鳥類的直系祖先。但是爬行動物和鳥類,恰好分屬於外溫性動物和內溫性動物。
由於這種複雜的關係,關於恐龍是否是內溫性動物,一直有廣泛的爭議。
 霸王龍會不會覺得熱?
霸王龍會不會覺得熱?古生物學家們嘗試了非常多的辦法去解決這個問題。但誰都不能帶著溫度計穿越回六千六百萬年前,找一隻霸王龍測一下(找到它估計也不會很配合)。所以之前,科學家們主要是通過恐龍的其他生理特徵和形態結構,側面研究恐龍是不是內溫性動物。
隨著研究深入,很多證據都支持恐龍可能具有中溫性,乃至內溫性。但由於缺乏直接的數據,對恐龍溫度調節狀況的描述,一直處於一個很模糊的狀態。
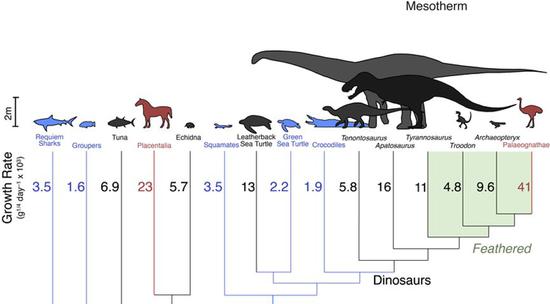 通過對恐龍和其他動物的生長速率這個間接證據,科學家們認為恐龍的體溫代謝情況接近中溫性動物
通過對恐龍和其他動物的生長速率這個間接證據,科學家們認為恐龍的體溫代謝情況接近中溫性動物穿越時光的“溫度計”
其實,恐龍的體溫是可以測量的。
科學家依靠的並不是月光寶盒、時間機器什麼的,而是δ47碳氧同位素團簇溫度計。
你一定想問:“還有這種可以穿越時光的溫度計?!”
其實這不是一種實體溫度計,而是一種地球化學研究方法。δ47碳氧同位素團簇溫度計是近些年來新興的恢復古溫度的地球化學手段,在古環境變化方面已經應用非常廣泛了。應用到古生物學當中的並不多,而用到恐龍化石當中更是稀少。
介紹這種方法,我們就要從這種方法的前身,氧18同位素溫度計開始說起。
說起同位素,大多數人一定會和“放射性”這個聽起來不太安全的名詞掛鉤。實際上,同位素是指具有相同質子數、不同中子數的同一元素的不同核素。同位素分為穩定同位素和放射性同位素。任何元素及其同位素都有固定的半衰期,放射性同位素是指那些半衰期較短(<1050a)原子核不穩定的同位素,如著名的鈾235。穩定同位素則往往相反,具有穩定的結構和長半衰期,相互之間往往只體現出質量數上的差異,由此導致物理、化學性質具有極其輕微的差異。
氧18同位素,是一種穩定同位素,由8個質子和10個中子構成。它們的數量相對我們熟悉的氧16要少很多,豐度為大約0.21%。氧18像氧16一樣穩定,在大多數情況下,它們除了稍微重一點,並沒有什麼異常,正常地存在水和空氣中。
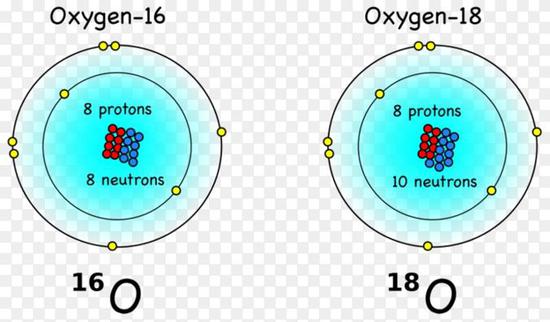 氧元素的兩種穩定同位素,氧16和氧18(圖片來源:維基百科)
氧元素的兩種穩定同位素,氧16和氧18(圖片來源:維基百科)某元素的同位素在物理、化學、生物等反應過程中,會以不同比例分配於不同物質之中,我們將這種現象稱為同位素分餾。我們用氧同位素舉個例子,水分子中含有氧元素,在水蒸發的時候,更多的含有更輕的氧同位素的水分子進入水蒸氣中,就像瘦子和胖子賽跑,瘦子跑的更快。
而不同溫度條件下,輕的和重的同位素“跑”的速度又不相同。基於這種原理,如果我們知道了某個環境下輕的同位素和重的同位素分佈的差異,也就是分餾的結果,就可以反推出這個環境的溫度情況了。基於這種巧妙的辦法,古環境學家們復原了地質歷史時期的溫度變化,我們現在能夠熱議的全球變暖,某種意義上也要歸功於古環境學家們成功恢復了古代的環境溫度。
威力加強版溫度計測到恐龍牙齒溫度
看到隔壁古環境學家們取得如此成就,古生物學家們肯定不甘示弱。這種可以測古代溫度的溫度計,為什麼不拿來測一測古生物化石呢?而後,古生物學家們也開始使用氧18同位素溫度計對化石進行測溫,但經過幾輪研究發現……
好像不是很好用啊!
這裡最大的問題不是測不出來,而是不知道測出來的到底是什麼的溫度。
動物體內氧元素的來源就是呼吸作用和飲用水。但不同溫度、不同地區,不同高度的氧同位素組成本身就有差異。動物喝的水里面氧同位素比例也有區別。還要考慮相當多的植食性動物攝入水分來自進食的植物,植物本身又對氧同位素有自己的分餾作用。
 作為陸地生物,恐龍身體裡的氧18同位素來源過於復雜並具有很強的環境依賴性。海拔、植被、距離海洋遠近等因素都能影響氧18同位素的含量(圖片來源:時代周刊)
作為陸地生物,恐龍身體裡的氧18同位素來源過於復雜並具有很強的環境依賴性。海拔、植被、距離海洋遠近等因素都能影響氧18同位素的含量(圖片來源:時代周刊)所以,雖然有一些基於氧同位素溫度計還原古生物體溫的研究,甚至恐龍當中也有涉及,但由於上述不確定性,一直不能算直接證據。雖然有些結果,但未被廣泛承認。
幸運的是,δ47碳氧同位素團簇溫度計技術的發展,讓古生物學家看到的新的曙光。簡單來說,這種溫度計是傳統氧18溫度計的“威力加強版”。δ47碳氧同位素團簇溫度計是基於碳的重同位素碳13和氧的重同位素氧18之間結合的程度,來推斷碳氧化學鍵生成的時候化學反應周遭的溫度。也就是說,可以準確測算礦物形成時,四周“小區域”的溫度,而很少受到大環境影響。
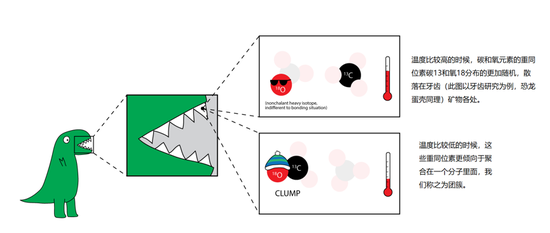 恐龍化石中應用δ47碳氧同位素團簇溫度計原理概述
恐龍化石中應用δ47碳氧同位素團簇溫度計原理概述古生物學是地質學和生物學的交叉學科,生物學上的概念給了這種新方法更廣闊的空間施展拳腳,那就是—— 內環境。
正如前文提到的,內溫性動物可以維持內環境溫度相對穩定在高於環境溫度的一個值附近。而氧18同位素溫度計的缺陷恰好就是環境因素對結果影響太大,說不清楚測到的溫度是“體溫”、“環境溫度”還是兩者混合。δ47碳氧同位素團簇溫度計恰好解決了這個痛點,測出的溫度是碳酸鹽礦物形成時周圍小環境的溫度,這樣理論上說就可以測定化石體溫了。
對於古生物學家來說,說這種方法是革命性的也一點也不過分!加州大學洛杉磯分校的環境地球化學學者Robert Eagle在2011年,將這種方法應用到了恐龍牙齒化石的牙釉質當中,通過牙齒化石中的碳酸鹽δ47碳氧同位素團簇溫度計來推斷恐龍體溫。
從牙齒到恐龍蛋到底哪個才是恐龍的體溫?
Robert Eagle的研究論文一經發表就引起了極大的關注。很多生物學家從生物的角度對這個工作提出了疑問:“牙齒的溫度,真的代表這種動物的體溫麼?”
通過研究現生動物,尤其是大體型動物,科學家們發現,很多動物都存在一個叫做“熱鑲嵌”(thermal mosaics)的生理現象。簡單來說,動物的體溫並不是均勻分佈的,可能只是軀幹的核心位置溫度較高而恆定,而身體邊邊角角的位置體溫可能相對更低。而牙齒,並不存在於恐龍身體軀幹的核心位置(恰恰相反,蜥腳類恐龍脖子很長,長在嘴裡的牙齒離軀幹更遠),因此測得的牙齒溫度,只能說是口腔溫度,不能代表恐龍的體溫。
 這個短吻鱷的紅外熱成像顯示,同一時刻,它的吻部溫度是明顯低於軀幹的。也就是說,它的溫度分佈模式是“鑲嵌”的
這個短吻鱷的紅外熱成像顯示,同一時刻,它的吻部溫度是明顯低於軀幹的。也就是說,它的溫度分佈模式是“鑲嵌”的我們知道,恐龍化石保存的都是骨頭,而恐龍體腔核心部分,也就是恐龍的內臟,是沒有化石保存下來的。如果用恐龍的骨骼化石做δ47碳氧同位素團簇溫度計體溫恢復,得出的結果會被永遠質疑不代表恐龍“血肉”的溫度。那怎麼辦呢?畢竟恐龍保存下來的都是骨頭,血肉是不能保存的。
不知道是Robert Eagle團隊靈機一動,還是受到高人指點。他們將實驗材料轉向了另一種化石材料——恐龍蛋。
為什麼是恐龍蛋呢?
我們用雞(也是一種現生的恐龍)生蛋的過程來還原一下。
母雞的卵巢裡有大約4000個初級卵母細胞,起先卵黃物質沉積在卵母細胞周圍,然後卵母細胞進入輸卵管。卵黃在輸卵管的漏斗部與精子結合,形成受精卵。受精卵繼續下行,在輸卵管的膨大部被蛋白包裹,形成蛋清。接著形成蛋殼膜,然後進入子宮,最後鈣質被沉積到蛋殼膜上,形成蛋殼以及蛋殼外面薄薄的角質層保護膜。
也就是說,對於蛋殼而言,蛋殼本身的碳酸鈣是在子宮附近形成並沉積的。而包括子宮、輸卵管在內的雌性生殖系統,一般都存在於雌性動物體腔內最核心的位置。只要測到了蛋殼的溫度,就能夠推斷出動物內環境的溫度。
不得不稱讚,這想法真是天才!
實驗流程基本不變,只需將分析材料換成更常見的恐龍蛋蛋殼化石。Robert Eagle團隊在2015年又報導了兩類恐龍的體溫。他們的研究發現,巨型長頸的蜥腳類恐龍體溫更高,在35°C到38°C之間,體溫可以與大部分現生的內溫性哺乳動物相當,略低於體溫更高的鳥類。而獸腳類恐龍的體溫要低一些,在32°C左右,在外溫性動物體溫的範圍內,比典型的內溫性動物還是要低一些。也就是說,小體型的獸腳類恐龍體溫低,而大體型的蜥腳類恐龍體溫高。這似乎印證了之前一些學者對恐龍體溫調節方式的判斷——巨溫性(簡單來說就是因為體型太大,散熱能力較低,所以體溫較高)。
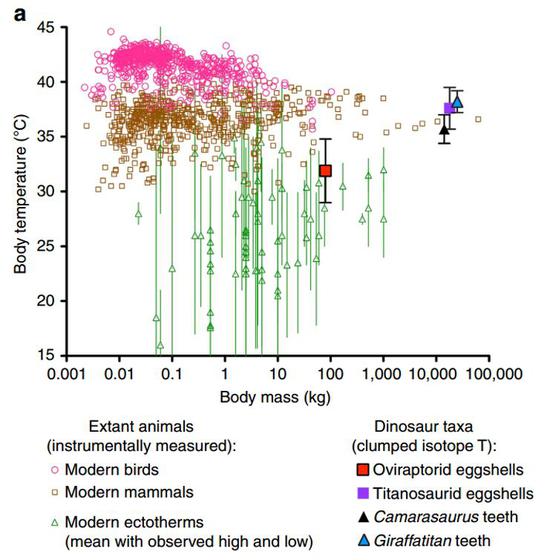 Robert Eagle的研究結果顯示,竊蛋龍的體溫在現生外溫性動物範圍內,而蜥腳類恐龍的體溫接近現生哺乳動物
Robert Eagle的研究結果顯示,竊蛋龍的體溫在現生外溫性動物範圍內,而蜥腳類恐龍的體溫接近現生哺乳動物需要解決的三個新問題
新的研究解決了舊的問題,但很快就又湧現出了新的問題。
首先是生物學層面的問題。不得不說的是,雖然這個研究實驗設計的很巧妙,但是取樣實在是太少了。雖然恐龍和恐龍蛋化石稀少,但Robert Eagle團隊兩次研究的採樣甚至都不能代表“非鳥恐龍”的全體。在恐龍的分類學框架下,一個重要的演化支——鳥臀類恐龍,沒有被納入到這個研究當中。
還有一個基本但很容易被忽視的問題。那就是在測量體溫之後,討論一種生物是否是內溫動物的時候,關注的是“保存體溫高於環境溫度的能力”,而不是“體溫的絕對值”。也就是說,理想情況下,我們應該看到測量出來的恐龍體溫,以及環境溫度,計算兩者的差距,如果差距非常明顯,才能真正證明恐龍是內溫性動物。Robert Eagle團隊的兩次研究,研究的化石都來自白堊紀的中低緯度地區,白堊紀的環境溫度本來就偏高,測量出來的較高的溫度可能不是因為這些恐龍是內溫性動物,而是因為環境很熱。這種情況在現生動物中是有例可循的,一些飼養在恆溫溫室的蜥蜴和龜(外溫性動物)體溫也可以很高,但是這不是它們自己調節控制的,而是依賴於外界環境的。
另外,一些來自地質學領域的問題也尚待解決。生物體死後形成化石,需要經歷漫長的埋藏和成岩作用。剛剛產下的蛋殼記錄了母體核心溫度,但經歷了超過七千萬埋藏的蛋殼是否還可以呢?實際上,如果化石在埋藏和成岩過程中經歷了重結晶作用,重新結晶形成新的碳酸鹽,那這些碳酸鹽帶來的溫度信息就與母體無關了,而反應的是地質作用時期的溫度。
可能就是因為前路荊棘遍布,也可能因為實驗條件苛刻,δ47碳氧同位素團簇溫度計在恐龍當中的應用自此停滯了接近五年的時間。
大家期待的直接證據,終於到來
是金子總會發光的,2020年2月14日,來自耶魯大學和劍橋大學的地質學家Robin Dawson和古生物學家Daniel Field在科學進展(Science Advance)雜誌上發表了基於δ47碳氧同位素團簇溫度計方法,對恐龍內溫性這個問題的最新研究。
雖然基本的研究方法繼承自Robert Eagle團隊,並且研究樣品也使用的是恐龍蛋殼。但新的研究工作通過新的研究手段和對比方法,大部分解決了Robert Eagle團隊的研究中存在的不足,進一步擴寬了人類對恐龍體溫這個問題的認識。
對於恐龍蛋化石的成岩重結晶作用造成的影響,此次研究對涉及的恐龍蛋材料,進行了蛋殼形態學、元素示踪和陰極發光顯微鏡等研究手段,去評估蛋殼受到地質作用影響,通過反算消去地質作用影響。
研究也首次對鳥臀類恐龍的代表慈母龍的恐龍蛋進行了δ47碳氧同位素團簇溫度計測溫工作。另外,為了避免低緯度地區研究材料不能證明內溫性,此次研究的材料大多來自高緯度地區(加拿大和羅馬尼亞),研究還同時對同地層的軟體動物和植物用同樣的方法測溫,來代表環境溫度,進而求得環境溫度和體溫數據的差值。
另外,此次研究的蜥腳類恐龍蛋殼來自羅馬尼亞的侏儒蜥腳類,是一種經歷了島嶼小型化的小型蜥腳類。這樣的體溫研究結果,也可以用來討論前面提到的“恐龍是否是巨溫性”的問題。
在Robin Dawson這篇文章的估計下:加拿大的獸腳類恐龍——傷齒龍的三個蛋殼樣品測出的體溫差距比較大,分別是38°C,27°C和28°C,但都顯著高於環境溫度,作者認為這代表了小型獸腳類恐龍可能是有一定的體溫變化範圍的,但也有相當高的體溫自我調節能力。
鳥臀類恐龍慈母龍的體溫就要高很多了,無論是否考慮成岩作用影響,它們的體溫估計都在40°C以上,最高估計可到44°C,可真是“發高燒”啊!相對而言,來自羅馬尼亞的侏儒蜥腳類體溫雖然較高,但沒有這麼極端,大約在36°C上下,和人的體溫非常接近。
同時,Robin Dawson發現,羅馬尼亞的侏儒蜥腳類的體溫測算結果和之前Robert Eagle團隊對體型更大的蜥腳類恐龍測算的體溫結果差不太多,侏儒蜥腳類和大型蜥腳類體重差了十倍,而體溫卻如此接近,因此她認為蜥腳類恐龍不具有“巨溫性”。最起碼可以認為在蜥腳類恐龍當中,體型和體溫之間關係並不大。
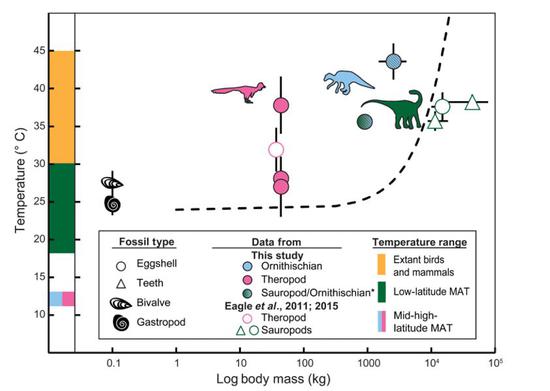 目前已經完成測溫的恐龍體溫和它們體型(x軸)以及生存環境溫度(y軸)之間的關係
目前已經完成測溫的恐龍體溫和它們體型(x軸)以及生存環境溫度(y軸)之間的關係縱覽現有的所有對恐龍各個類群(鳥臀類,蜥腳類和獸腳類)代表的δ47碳氧同位素團簇溫度計體溫估計,我們可以發現,雖然恐龍內部當中體溫有所差異,但都是顯著的高於環境溫度的。作者整體評估了恐龍的溫度調節能力,認為它們應該類似現生的中溫性動物,如棱皮龜。系統研究顯示這種能力存在於各個恐龍演化支系當中,也就意味著,這種相對主動和積極的體溫調節策略,可能自恐龍的祖先開始,就是恐龍家族的“傳家寶”。也許,這也是它們能夠統治中生代陸地的原因之一。
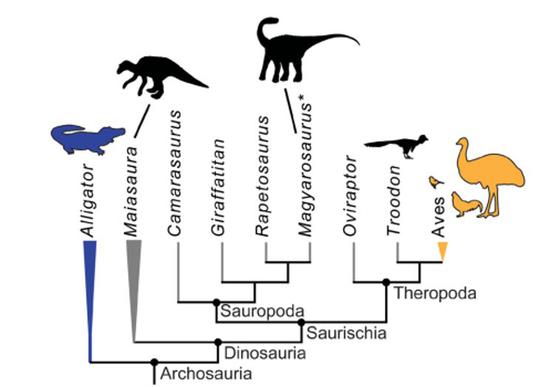 Figure 9 恐龍的後代是典型的內溫性動物,鳥類;而恐龍遠親鱷類則是典型的外溫性動物;目前對恐龍內部各大類群的測溫顯示,可能所有非鳥恐龍都具有維持體溫高於環境溫度的積極的體溫調節能力(圖片來源:Dawson et.al 2020)
Figure 9 恐龍的後代是典型的內溫性動物,鳥類;而恐龍遠親鱷類則是典型的外溫性動物;目前對恐龍內部各大類群的測溫顯示,可能所有非鳥恐龍都具有維持體溫高於環境溫度的積極的體溫調節能力(圖片來源:Dawson et.al 2020)